I – II – III – IV – V – VI – VII – VIII – IX – X – XI – XII – XIII – XIV – XV – XVI – XVII – XVIII – XIX – XX – XXI – XXII – XXIII – XXIV – XXV – XXVI – XXVII – XXVIII – XXIX – XXX – XXXI – XXXII – XXXIII – XXXIV – XXXV – XXXVI – XXXVII – XXXVIII – XXXIX – XL – XLI – XLII – XLIII – XLIV – XLV – XLVI
[Single-page view]
Speaking of cosmology, though, I’d be remiss if I didn’t talk about the most important biblical story of all – the Genesis creation story – and spend a section delving into the whole question of creationism versus evolution. According to the literal reading of the Bible, the universe (along with all the living creatures in it) was created over the course of six days, about 6,000 years ago. (This date was most famously calculated by Archbishop James Ussher in 1650, by adding up the ages and birthdates of all the biblical patriarchs, and tracing them all the way back from Jesus to Adam. Others have crunched these same numbers and come up with the same result.) But to say that this figure contradicts everything science has been able to determine about the age of the universe is an understatement, to say the least.
All of the scientific evidence shows that the universe is, in fact, about 13.8 billion years old, that the earth itself is about 4.6 billion years old, that life first began to emerge about 4 billion years ago, that the first hominins appeared about 7 million years ago, and that modern Homo sapiens first appeared around 300,000 years ago. It also shows that this process was anything but a quick six-day event; the way we actually got here was through a series of extremely long and gradual physical processes, including the process of biological evolution.
The concept of evolution can be a hard one to swallow for a lot of religious believers, not just because their religions require them to reject the idea, but just because intuitively it seems too strange to imagine that one type of creature could turn into a completely different type of creature given enough time. Still, even the most ardent creationists acknowledge the reality of, for instance, dog breeding – the fact that ancient humans domesticated wild wolves, selected the ones with the most desirable traits, and continued doing that for generation after generation until eventually they ended up with Dalmatians and Chihuahuas and Saint Bernards and so on. And really, that’s all evolution is. We all recognize the reality that parent organisms pass along physical traits to their offspring – that’s why children tend to look like their parents. We also recognize that sometimes there are variations in these inherited traits – that’s why children of the same parents don’t all look or act exactly alike. And we recognize that through processes like selective breeding, certain traits can become exaggerated in later generations – that’s why greyhound breeders have been able to produce faster greyhounds over the years, why bulldog breeders have been able to produce squattier bulldogs, and so forth. Human intervention isn’t the only factor that can affect a species’ breeding pool, of course; as QualiaSoup points out, “If a greyhound breeder selects only the fastest dogs for breeding stock, and in the wild only the fastest gazelles outrun their predators and survive to reproduce, then both nature and the greyhound breeder are favoring certain individuals for reproducing offspring and passing on their genetic information to the next generation.” (His video below is a must-watch.) But once you accept all of these self-evidently true statements, then that’s it – you’ve accepted evolution. “Descent with modification” is literally all evolution is. True, the variations between one generation and the next will always be relatively minor in the short term – a wild wolf will never give birth to a dachshund, just like a wild ape will never give birth to a human – but over millions of years (or less if the evolution is human-guided), tiny incremental changes can and do add up; and with enough time, you can go from one primitive parent species to an extraordinarily rich and diverse family tree. You can even go from simple microscopic organisms to all the complex biological diversity we see today. And as all the evidence from every relevant scientific field shows us, this is exactly what has been happening on Earth over the last 4 billion years.
Some of the best evidence for evolution comes from instances in which we can actually see the evolutionary process happening directly. The classic example always given in biology classrooms (and briefly alluded to in QualiaSoup’s video) is that of the peppered moth, which was common in Britain around the time of the Industrial Revolution. As the Butterfly Conservation website explains:
Peppered Moths are normally white with black speckles across the wings, giving it its name. This patterning makes it well camouflaged against lichen-covered tree trunks when it rests on them during the day. There is also a naturally occurring genetic mutation which causes some moths to have almost black wings. These black forms (called ‘melanic’) are not as well camouflaged on the lichen as normal ‘peppered’ forms and so they are more likely to be eaten by birds and other predators. This means that fewer black forms survive to breed and so they are less common in the population than the paler peppered forms. This is the normal situation observed in the countryside of Britain and Ireland.
However, in the nineteenth century it was noticed that in towns and cities it was actually the black form of the moth that was more common than the pale peppered form. Industrialisation and domestic coal fires had caused sooty air pollution which had killed off lichens and blackened urban tree trunks and walls. So now it was the pale form of the moth that was more obvious to predators, while the melanic form was better camouflaged and more likely to survive and produce offspring. As a result, over successive generations, the black moths came to outnumber the pale forms in our towns and cities. Since moths are short-lived, this evolution by natural selection happened quite quickly. For example, the first black Peppered Moth was recorded in Manchester in 1848 and by 1895 98% of Peppered Moths in the city were black.
In the mid-twentieth century controls were introduced to reduce air pollution and as the air quality improved tree trunks became cleaner and lichen growth increased. Once again the normal pale Peppered Moths were camouflaged and the black forms were more noticeable. Now the situation in urban areas has again become the same as in the countryside, with normal pale Peppered Moths being far more common than the black forms. So natural selection has been seen to work in both directions, always favouring the moth that is best suited to the environmental conditions. The same thing has been observed throughout Europe and the USA.

Another good piece of evidence that we can directly observe for ourselves is the existence of “ring species.” Occasionally, a particular species will spread across such a large geographic area that the population at one end of the distribution will find itself in a completely different habitat from the population at the other end, and will accordingly evolve different traits; then, when the different populations continue spreading until they eventually converge again, they’ll have become so different by that point that they’ll be reproductively isolated from each other and will no longer interbreed. They’ll have evolved into two different species, despite still being linked by a continuous chain of populations of the same species that they can and do still interbreed with. In other words, they’ll represent a real-life branching of the evolutionary tree. Potholer54 explains:
Creationists will often concede that evolution does happen on the “micro” scale within species, but then deny that it can create new species on the “macro” scale. But the example of ring species shows just how illusory this distinction actually is. All macroevolution is, really, is just a bunch of accumulated microevolutions.
And there are still more ways in which we can see the branching of the evolutionary tree directly. In short-lived species like insects and bacteria, their rapid reproductive cycles allow them to accumulate so many changes over such a short period that we can actually see them evolving in real time (as opposed to larger species, where evolution typically takes millions of years). Scientists have been able to observe insect speciation in laboratories (i.e. one population of interbreeding insects evolving into two populations with different characteristics that can no longer interbreed) on dozens of occasions; and you could even recreate their results yourself if you really had a mind to. Then again, you’ve probably already had quite a few firsthand encounters with evolution in rapidly-reproducing species; after all, it’s the reason why you have to get a new flu shot every year. The strain of influenza that threatens to make you sick in any given year never stays the same – it’s always evolving – so by the time the next year rolls around, the old shots don’t work on it anymore, and you need to get a newer version.
To take another example of ultra-rapid microbial evolution: There was a bacterium discovered in 1975 that subsisted on manmade nylon byproducts – something no other bacterium was capable of doing. This might not have seemed particularly remarkable, except for the fact that nylon is completely synthetic and wasn’t even invented until 1935 – so the only way this bacterium could have come into existence is if it had evolved sometime after that date. Sure enough, when scientists tested the theory by putting a totally different species of bacterium in the same kind of nylon-rich environment with no other source of nutrients, it evolved the same mutation. And using similar techniques, they’ve been able to observe the process of speciation in other kinds of bacteria as well, as VoysovReason discusses:
Another well-known study is the long-term evolution experiment led by Dr. Richard Lenski, using E. coli bacteria. Since bacteria reproduce very quickly, they provide an accelerated way to observe evolution. Since 1988 – and continuing to this day – Lenski has been tracking genetic changes in 12 initially identical populations of E. coli. As of 2014, each of the 12 isolated populations have reproduced for 58,000 generations. The researchers have observed a wide array of genetic changes. Some evolutionary adaptations have occurred in all 12 populations, while others have only appeared in one or a few populations. One particularly striking adaptation was the evolution of a strain of E. coli that was able to use citric acid as a carbon source in an oxygen-rich environment – a completely new trait. This clearly demonstrates how evolution produces new species. Of course, the descendants of the bacteria are still bacteria, as creationists like to point out – but they’re a new kind of bacteria; and that is how evolution always works. Mammals are a special kind of tetrapod. Primates are a special kind of mammal. Apes are a special kind of primate. And humans are a special kind of ape. Animals never entirely stop being what their ancestors were; they just change into new versions of that animal. It generally takes large spans of time before the differences outnumber the similarities and the new animals are considered a new species.
This last point is underscored by the fact that we can so often see examples of species that are still in the middle of a particularly conspicuous evolutionary transition, defying the idea of well-defined biblical “kinds.” For instance, the most iconic image that pops into everyone’s heads when they imagine “evolution in action” is that of a prehistoric fish species gradually gaining the ability to crawl onto land and breathe air. It’s an idea that creationists will sometimes scoff at – but the thing is, there are actually species of fish alive today, like lungfish and mudskippers, that can crawl onto land and breathe air. If we’re going to deny that any kind of transition between fish and land creatures is even possible, then what are we to make of these species?
Similarly, if we take seriously the biblical idea that lizards naturally have legs but snakes don’t (since God cursed the serpent in the Garden of Eden), then what are we to make of the lizard species existing today whose legs are in the process of growing progressively smaller (beyond the point of being functional) and in some cases have disappeared entirely? Or for that matter, what about the snake species that still retain tiny nubs where their legs haven’t completely finished disappearing yet?

It isn’t just a handful of species that raise these kinds of questions, mind you – every species displays some remnant of its earlier evolutionary stages within its modern-day anatomy. It’s just that certain cases (like snake legs) are more obvious than others because the anatomical remnants in question are no longer serving a useful purpose – they’re just leftover redundancies, which biologists refer to as vestigial features.
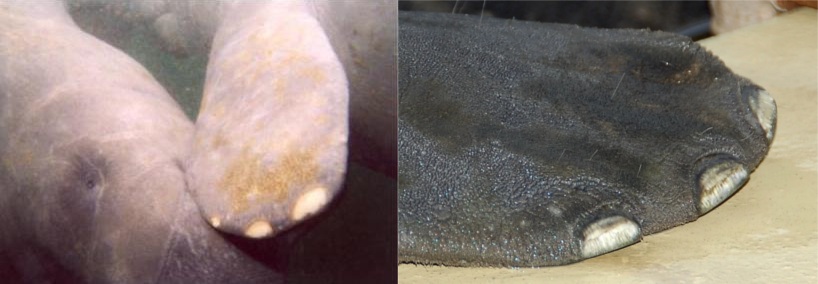
Some of the best examples of vestigiality can be found in aquatic mammals like whales, dolphins, and manatees, which originally evolved from four-legged land-dwelling species. Like snakes, they have rudimentary pelvises and hind leg bones that have been so reduced from their ancestral form that they can no longer be used for their original function. (The pelvic bones, at least, do still serve as anchors for the muscles of the sex organs; but there’s no functional purpose for the femur bones connecting to them via ball-and-socket joints.)

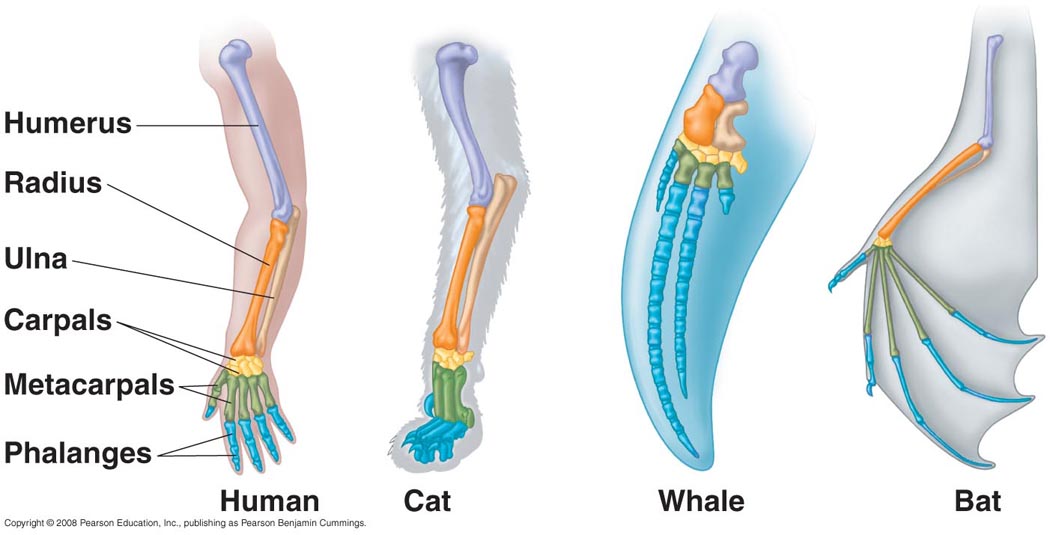
Their forelimbs also show evidence of their ancestry; instead of having paddle-like fin bones (like fish and sharks do), the bones forming their fins take the form of “hands” that have simply been repurposed for swimming – with each forelimb having a clearly defined proximal bone (humerus), two distal bones (radius and ulna), carpals (wrist bones), metacarpals (palm bones) and phalanges (fingers). You might easily mistake the image below for an ape’s hand, but it actually belongs to a dolphin:

In fact, this basic skeletal structure is shared by all classes of tetrapods (from amphibians to mammals) with varying degrees of modification, since they all descended from the same common ancestor. They’ve all just evolved variations on this common skeletal blueprint to fit their different evolutionary niches; so what started off as one basic ancestral limb has evolved to serve all kinds of different functions in different descendent species.
This also explains why manatees can often be found with elephant-like toenails at the end of their flippers – they’re evolutionary leftovers from a time when the manatees’ land-dwelling ancestors actually had toes.
And evolution accounts for other such features as well, like the fact that whales still have trace amounts of body hair despite having no use for it – or the fact that, for that matter, they still have lungs rather than gills and have to come up for air every half hour or so (or in the case of dolphins, every ten minutes) despite living their whole lives underwater. (Sad fact: When a whale or dolphin dies of “natural causes,” it’s because it has gotten so old that it no longer has the strength to resurface and simply runs out of oxygen.)

We don’t have to stop at manatees and whales, though; vestigiality can be found everywhere. There are blind rodents that still have eyes, but their eyes no longer function and are covered with a layer of skin, because the rodents live underground and have no need for sight. Evolution is still in the middle of a millennia-long process of eliminating their eyes; it just hasn’t finished with them yet. (There are also cave-dwelling fish and salamanders undergoing the same process themselves.) Similarly, there are flightless beetles that still retain wings beneath their shells that they are incapable of using; and there are flightless birds with wings that have likewise ceased to serve any valuable function. Many of these flightless birds also have the added trouble of dealing with bones that are still hollow – perfect for flight, but a liability on the ground where they’re more likely to break. And in bats, the reverse is true – they still retain the solid bones of their land-dwelling ancestors, so flight is more difficult than it needs to be. In all of these cases, such “design flaws” only make sense in the context of evolution. These animals weren’t deliberately designed with these flaws at all; they just inherited them.
One of the most glaring examples worth mentioning here is the recurrent laryngeal nerve of the giraffe. This nerve, found in all vertebrate species with necks, connects the brain to the larynx – a trip that, in theory, should only take a few inches. But due to a weird evolutionary fluke, the nerve instead travels all the way down the giraffe’s neck, loops around the aortic arch of the heart, and then travels all the way back up the neck to reach its destination – an unnecessary detour of about 15 feet. As the Wikipedia article explains:
The nerve’s route would have been direct in the fish-like ancestors of modern tetrapods, traveling from the brain, past the heart, to the gills (as it does in modern fish). Over the course of evolution, as the neck extended and the heart became lower in the body, the laryngeal nerve was caught on the wrong side of the heart. Natural selection gradually lengthened the nerve by tiny increments to accommodate, resulting in the circuitous route now observed.
This particular design flaw is one that shows up in humans as well (albeit not quite as dramatically as with giraffes). And it’s not the only one. I’ve only been talking about non-human animals up to this point, but it’s also worth talking about some examples where evolution has left its mark on our own species. Ever wonder, for instance, why we get goosebumps when we’re cold or scared? As it turns out, this is a vestigial leftover from a time when our ancestors were covered in thick coats of hair, and getting goosebumps was a way to raise those hairs and “fluff themselves up” to keep themselves warm in the winter or make themselves appear larger and scare off predators. Similarly, we also have muscles connected to our ears which, eons ago, allowed our ancestors to swivel their ears around like antennae (in the same way that horses and rabbits can), but which have since lost that function and are no longer useful. There’s also the fact that we grow wisdom teeth – which would have been useful in our larger-mouthed ancestors, but which now only cause unnecessary complications and often have to be painfully removed. There’s the fact that our legs, feet, and spines haven’t fully adapted yet to supporting the weight of an upright, two-legged posture – so that as we age we often experience hip and knee pain, lower back pain, collapsed arches in our feet, and other such issues that four-legged animals don’t typically have to deal with. There’s the fact that the male urinary tract runs directly through the prostate gland, so that when the prostate becomes swollen (as it does for most men later in life), it creates needless and sometimes dangerous urinary problems. There’s the fact that the female pelvis is too small to easily give birth to the large-headed newborns of our species – a feature that wouldn’t have been an issue in our smaller-headed ancestors, but which leads to immense suffering for mothers everywhere today (and in millions of cases, leads to complications and even death). There’s the fact that our respiratory and gastrointestinal tracts (i.e. the windpipe and the esophagus) share the same tube – so that we often accidentally choke to death – rather than using two separate tubes as they do in dolphins. There’s the fact that the blood vessels in our eyes run across the front of the retina rather than beneath it, leading in many cases to vision loss – as well as the fact that our photoreceptors are inefficiently oriented backwards, resulting in a blind spot in the middle of our vision (a flaw that’s absent in animals like octopuses and squid, whose photoreceptors are more sensibly oriented forwards). And the list goes on.
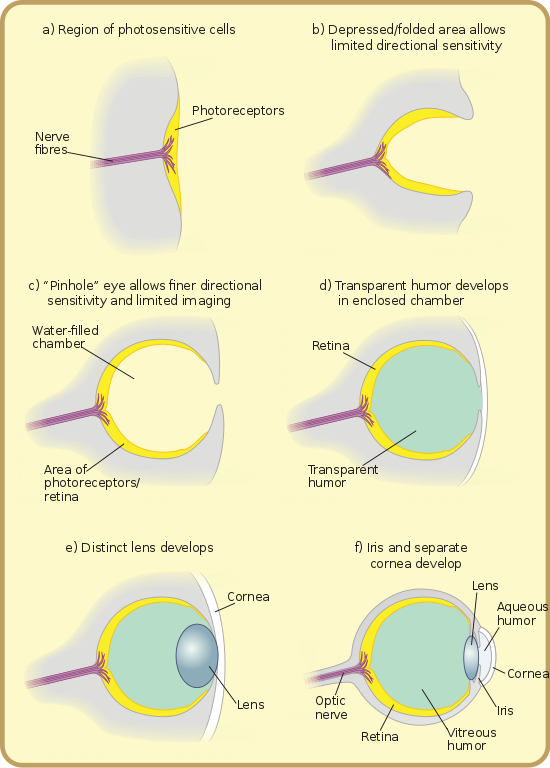
Speaking of eyes, there’s an argument that creationists often make against evolution, which asks, “how could something as complex as the eye have simply come into existence on its own?” But the answer is that the eye didn’t just pop up out of nowhere (something like that really would be too implausible to believe); rather, it developed very gradually, one small step at a time. Originally, the eye started off as nothing more than a patch of light-sensitive cells on some prehistoric organism (like the light-sensitive spot on a modern-day euglena). This patch of cells wouldn’t have allowed for true vision, of course, but it would have allowed the organism to distinguish between light and dark, giving it a small evolutionary edge and making it more likely to survive and pass on this feature to its offspring. Later on, some of this species’ descendants would be born with slight depressions in their light-sensitive patches, giving their primitive eyespots a cuplike shape and enabling them to distinguish not only whether it was light or dark, but which direction the light was coming from (like in the eyespots of a modern-day planarian). Generations after that, the opening of this cup would narrow to form a small aperture, like a pinhole camera, allowing light to enter and produce a dim, fuzzy image (like in the eyes of a nautilus). Later still, a transparent cover would form over the opening, allowing it to fill with fluid and sharpening the image resolution considerably (like in abalone eyes). And finally, the eye would assume the “finished” form that we currently see in mammals. Every step in this process was gradual – there were no sudden leaps between a primitive form and a much more complex one – but as the old saying goes, a journey of a thousand miles begins with a single step. And that’s how it works with evolution.
Returning to the topic of vestigiality, though, there’s another phenomenon that’s closely related and provides a similar kind of evidence for evolution – the existence of atavisms. In the same way that vestigial traits are anatomical features left over from earlier ancestors, atavisms are basically leftover genes. Underlings explains:
These are dead genes that occasionally mutate and switch back on, resulting in an individual growing a feature no longer expressed by typical members of the species. For example, one in 500 whales is born with external rear leg remnants, sometimes even possessing feet and toes. Horses have only one toe per leg, but occasionally some are born with up to three toes, just like their extinct relatives. Sometimes humans are born with functioning tails, just like monkeys. It’s even possible to cause chickens to develop teeth-like structures by providing a single missing protein to otherwise dead genes. These degenerating leftover genes are what we might expect to find if whales evolved from a land mammal, horses evolved from a three-toed ancestor, humans evolved from a tailed primate, and birds evolved from dinosaurs with teeth – but what sense do they make if all species were instead unique creations by a perfect God?

The creationist response to this question may be that, well, since the same God created birds and mammals and reptiles and everything else, it’s no surprise that he gave them all the same varieties of genes. The fact that dead genes sometimes get accidentally activated proves nothing about how organisms are related, because God used the same genetic blueprint for all of them anyway. But the thing is, atavisms only appear in organisms that are ancestrally related on the evolutionary tree; in other words, mammals and birds may both display atavisms from reptiles (because they both evolved from reptiles), but you’ll never see mammals displaying atavisms from birds, or vice-versa, because neither mammals nor birds evolved from each other. And it’s the same way with vestigial features, as the Cassiopeia Project explains:
There are no vestigial structures that were not previously functional in an ancestor. All vestigial organs make sense only in the framework of evolution; and of course we do not find vestigial organs that argue against evolution. No nipples in amphibians, or vestigial feathers on mammals; no primates carry vestigial horns or degenerate wings; we do not find arthropods with leftover backbones; no snakes have wing parts; and no humans have gizzards.
The patterns in which atavisms and vestigial features appear don’t just show that organisms are interrelated; they show how they’re related, and how they evolved from each other. They point to one very specific evolutionary tree – and evidence for this tree can be found in other fields as well, like embryology. Underlings continues (starting at the 10:45 mark):
That last line is blunt, but it raises a fair point. If we take it for granted that all life on Earth really was consciously designed by a perfect, all-knowing creator, then it’s exceptionally hard to explain why there are so many flaws and redundancies in the designs (aside from just hand-waving it away with the rationalization that God must have had his reasons). If we turn to the evolutionary explanation, though, it all makes perfect sense. And in fact, we don’t even have to wonder where all these imperfect anatomical features came from, because in most cases we can trace these species’ full family histories back through the fossil record and see exactly where they came from, how they evolved, and how long the process took.
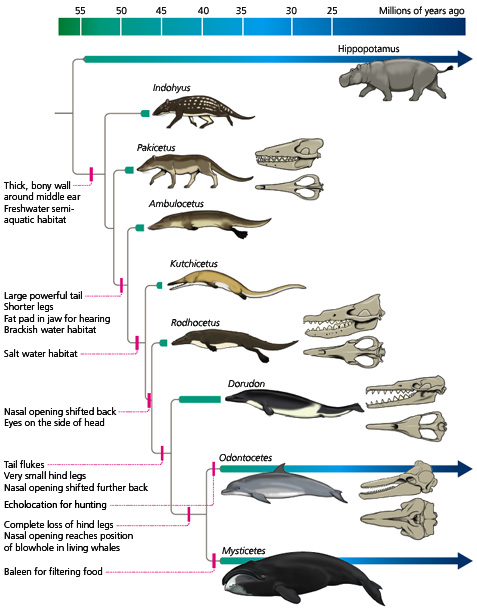
In the case of whales, for instance, we can examine fossils of their most immediate ancestors and see that they’re almost identical to their modern form. Then if we dig deeper into the fossil record, we can see that their ancestors slightly before that were still similar, but had more developed hind limbs, blowholes that were further toward the front of their skulls, and so on. Dig further still, and these differences become more and more pronounced, with the legs becoming larger, the nasal openings migrating closer to the front of the snout, etc. – until eventually you can trace the whales’ ancestry all the way back to hippo-like animals with four fully-developed legs rather than flippers, regular nostrils instead of blowholes, and all the other adaptations necessary for life on land.
Similarly, we can trace back the lineage of modern horses, from the large modern species with their single-hoofed feet, to their much smaller ancestors with multiple toes on each foot, in a smooth, continuous timeline of fossil specimens, each of which is just a slightly modified version of its predecessor.
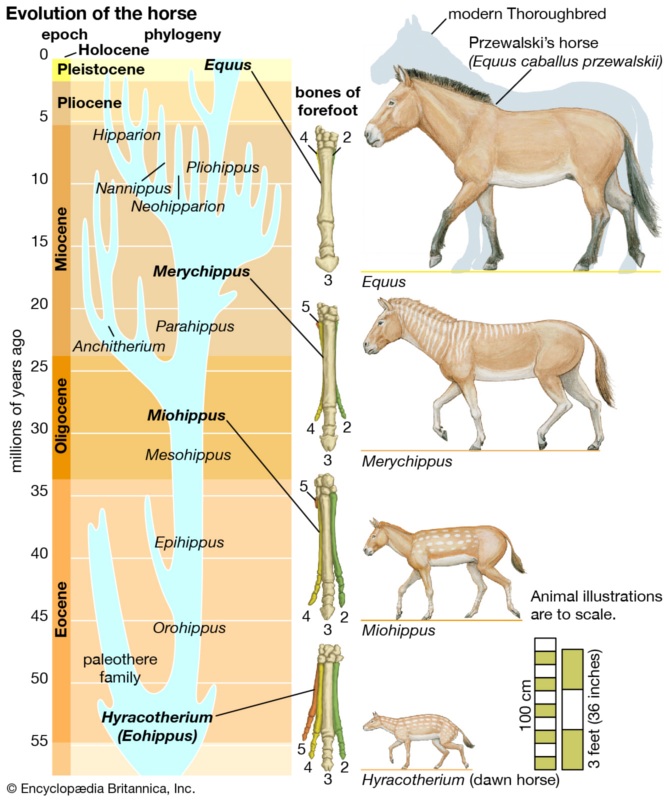
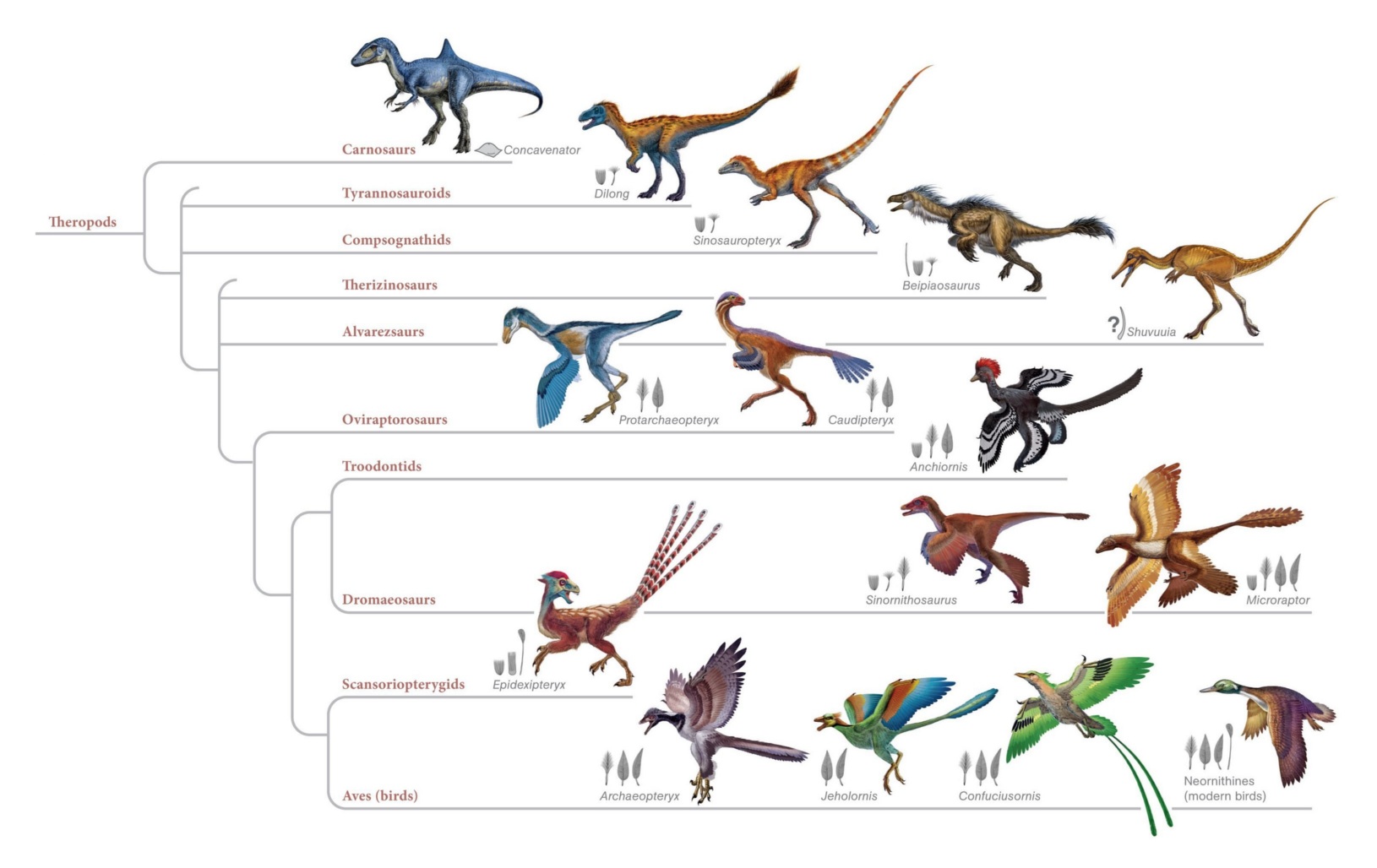
And we can do this with countless other species as well. Probably the most famous transitional fossil of all is Archaeopteryx, a creature halfway between a dinosaur and a modern-day bird. Discovered just two years after Darwin published his book On the Origin of Species and predicted that such transitional fossils ought to exist (even though none had yet been discovered), Archaeopteryx was notable for having the clawed fingers, bony tail, toothed jaws and snout of a theropod dinosaur, and the wings, flight feathers, and hollow bones (including a wishbone) of a bird. It was the perfect illustration of an evolutionary transition between one “kind” of animal and another; and as the years passed and more transitional fossils were discovered, the ancestral chain between dinosaurs and their bird descendants has – once again – been filled in with a smooth, continuous progression of intermediate steps.

A crucial point to mention here is that these progressions of transitional fossils always occur in a very specific geological order. The earth’s crust is made up of different strata (i.e. layers) of rock, with the oldest layers at the bottom and more recent layers on top (which makes sense, considering that the newer layers wouldn’t have been able to accumulate on top of anything if there weren’t already older layers beneath them). Scientists have confirmed that the lower layers are older using a variety of dating methods, which we’ll get into momentarily; and you can see the layers for yourself when you look at the Grand Canyon or some other geographic feature where there’s a natural cutaway view of the rock.
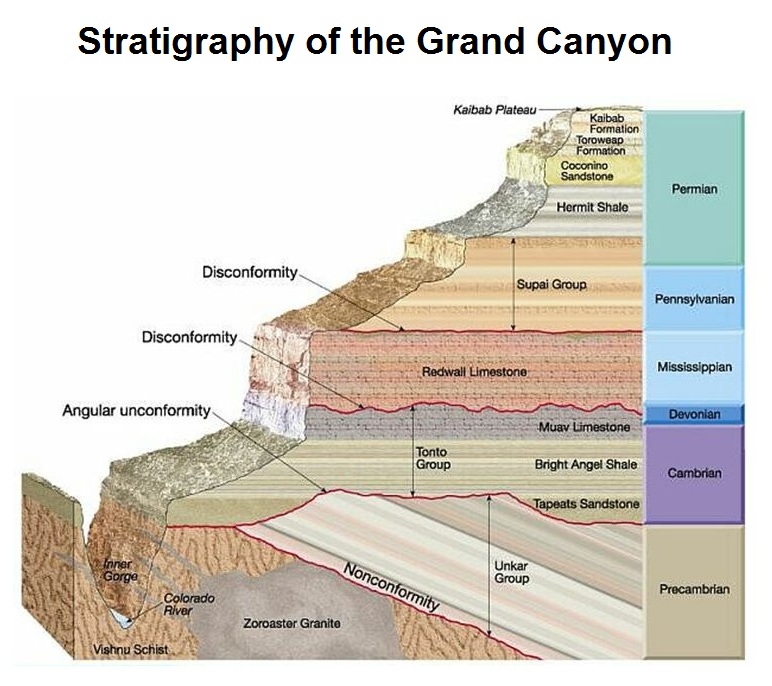
The relevance of this, in terms of evolution, is that when you excavate fossils from these different layers, the more primitive organisms are always found exclusively in the older layers, while the more recently-evolved organisms are always found exclusively in the younger layers. There are never any fossils of recently-evolved organisms found in the lowest strata of the geologic column, nor are there ever any fossils of the most primitive organisms found in the highest strata. If creationism were true, then this wouldn’t make any sense; we would see trilobite fossils, dinosaur fossils, and giraffe fossils all buried in the same layers of rock, since they were all created at once and they all lived together at the same time. But we never see that kind of simultaneity; it’s always the case that the more primitive an organism is, the older and deeper its fossils are.
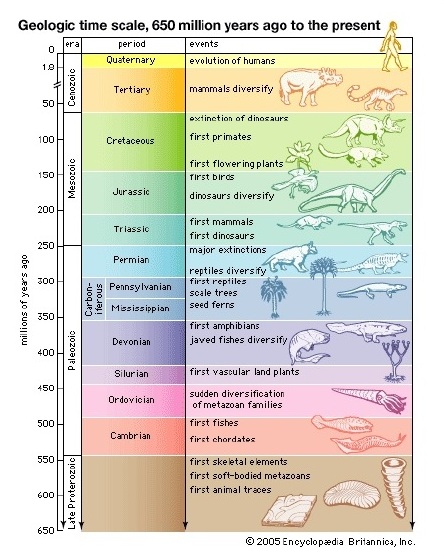
In fact, this pattern is so consistent that scientists have actually been able to use it to successfully predict where the fossils of new transitional species will be found and what they’ll look like. Underlings explains (from 2:21-3:31):
This kind of thing simply shouldn’t be possible if creationism were true. But like Underlings says, it has proven so reliable that we can now trace the evolutionary ancestry not only of all these various animal species, but even of our own species as well.
